Forschungsprojekte
Überblick
Ein fundamentales Prinzip von Zellen ist ihre Fähigkeit, funktionsfähige biologische Systeme an Veränderungen der Umgebung anzupassen. Dafür verfügen Zellen über Schutzmechanismen, die die Integrität und Funktionalität ihres Proteoms überwachen. Zelluläre Kontrollsysteme erkennen Fehlfunktionen oder Beschädigungen individueller Komponenten und lösen adaptive Stressantworten aus, um das Überleben sowohl auf zellulärer als auch auf Ebene des Gesamtorganismus durch das Arrangieren ihrer Reparatur, Entfernung und Austausch zu gewährleisten. Eine Fehlfunktion oder Deregulation dieser Proteinhomöostase wird für das Altern und verschiedene schwere Erkrankungen wie Krebs, Neurodegeneration und entzündliche Krankheiten verantwortlich gemacht. Wir möchten die Mechanismen verstehen, die die Proteinhomöostase in Zellorganellen kontrolliert.
Die Forschungen unserer Arbeitsgruppe fokussieren sich auf drei Hauptaspekte:
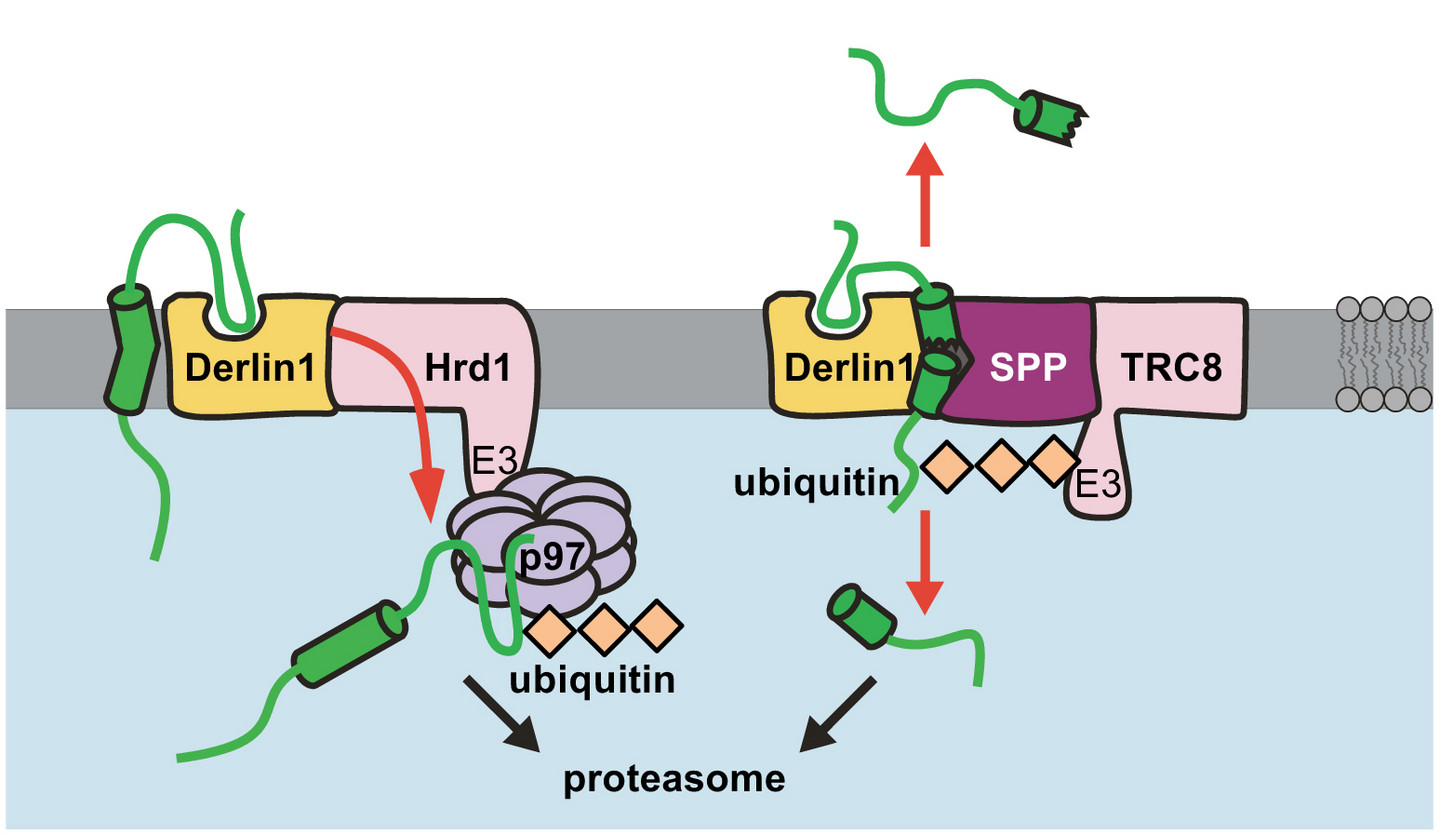
1.) Intramembranproteolyse – ein neuer regulatorischer Zweig des ERAD Abbauwegs
Ungefähr ein Drittel aller Säugerproteine wie beispielsweise wichtige zelluläre Oberflächenrezeptoren und Kanäle werden im Endoplasmatischen Retikulum (ER) synthetisiert. Der ER-assoziierte Abbauweg (ERAD) ist besonders wichtig, um zur Aufrechterhaltung der Proteinhomöostase missgefaltete und beschädigte Proteine zu entfernen. Allerdings kann auch die Menge nativer Proteine über ERAD kontrolliert werden. Die ERAD Maschinerie umfasst verschiedene parallele Wege, die die Erkennung und Entfernung eines heterogenen Substratspektrums ermöglichen. Wir und andere konnten zeigen, dass Intramembranproteasen und deren katalytisch inaktive Homologe, sogenannte Pseudoproteasen, als zentrale ERAD Faktoren die Menge und Aktivität selektiver Membranproteine kontrollieren. In unserem Labor benutzen wir eine Kombination zellbiologischer, biochemischer und proteinchemischer Techniken, um die physiologischen Funktionen und molekularen Mechanismen ER-residenter Proteasen und Pseudoproteasen bei der Proteinhomöostase zu untersuchen.
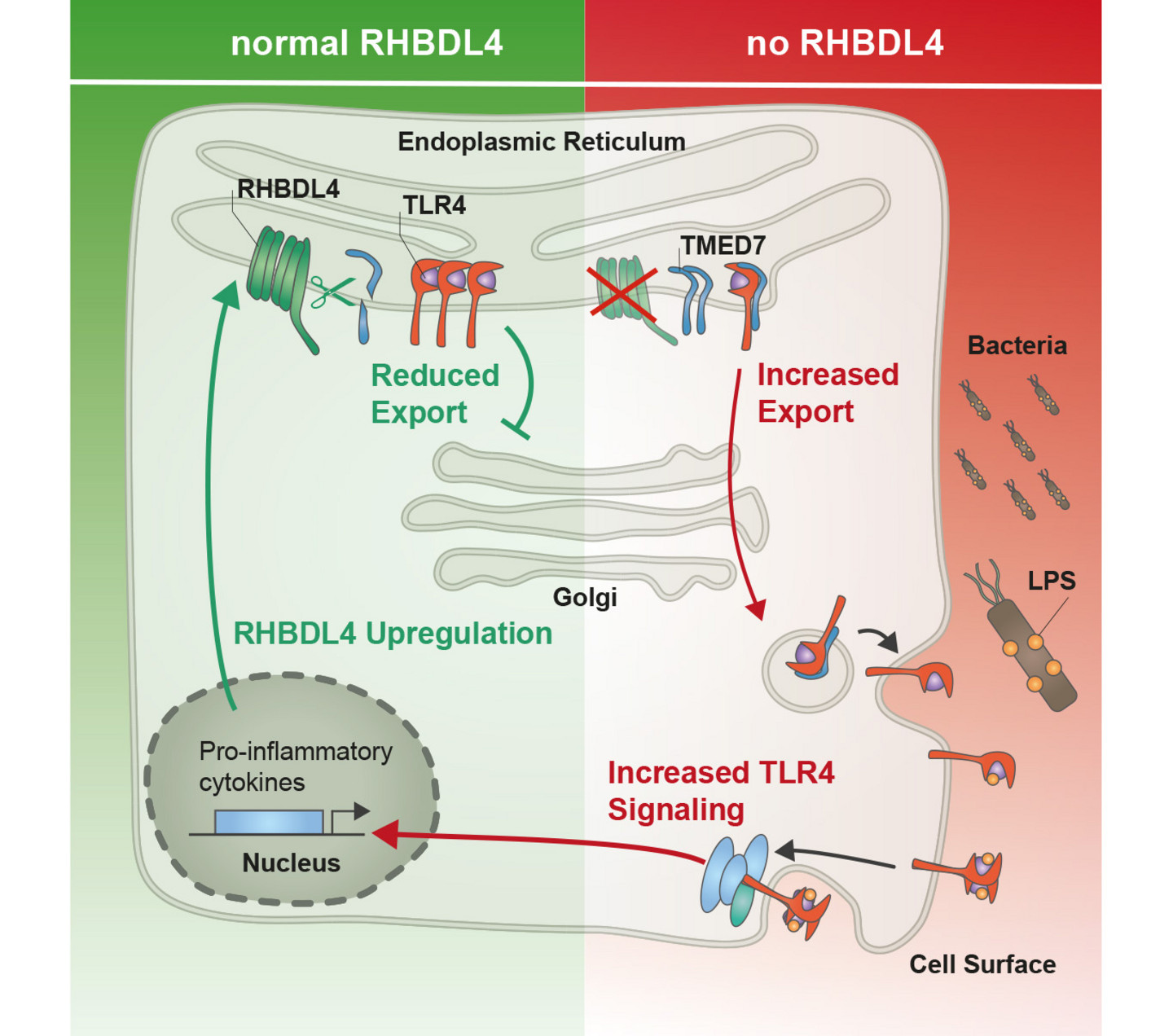
2.) Molekulare Mechanismen der regulierten Proteinsekretion
Korrekt gefaltete Proteine werden vom ER abhängig von spezifischen Frachtrezeptoren mittels sogenannter COPII-umhüllter Vesikel zum Golgi-Apparat geschleust. Wir wollen klären, wie die Frachtauswahl an den „ER-Exit Site“ reguliert wird. Kürzlich haben wir gezeigt, dass Aktivität von p24-Frachtrezeptoren durch Intramembranproteolyse reguliert und dadurch wichtige Plasmamembranproteine wie der Immunrezeptor TLR4 kontrolliert werden.
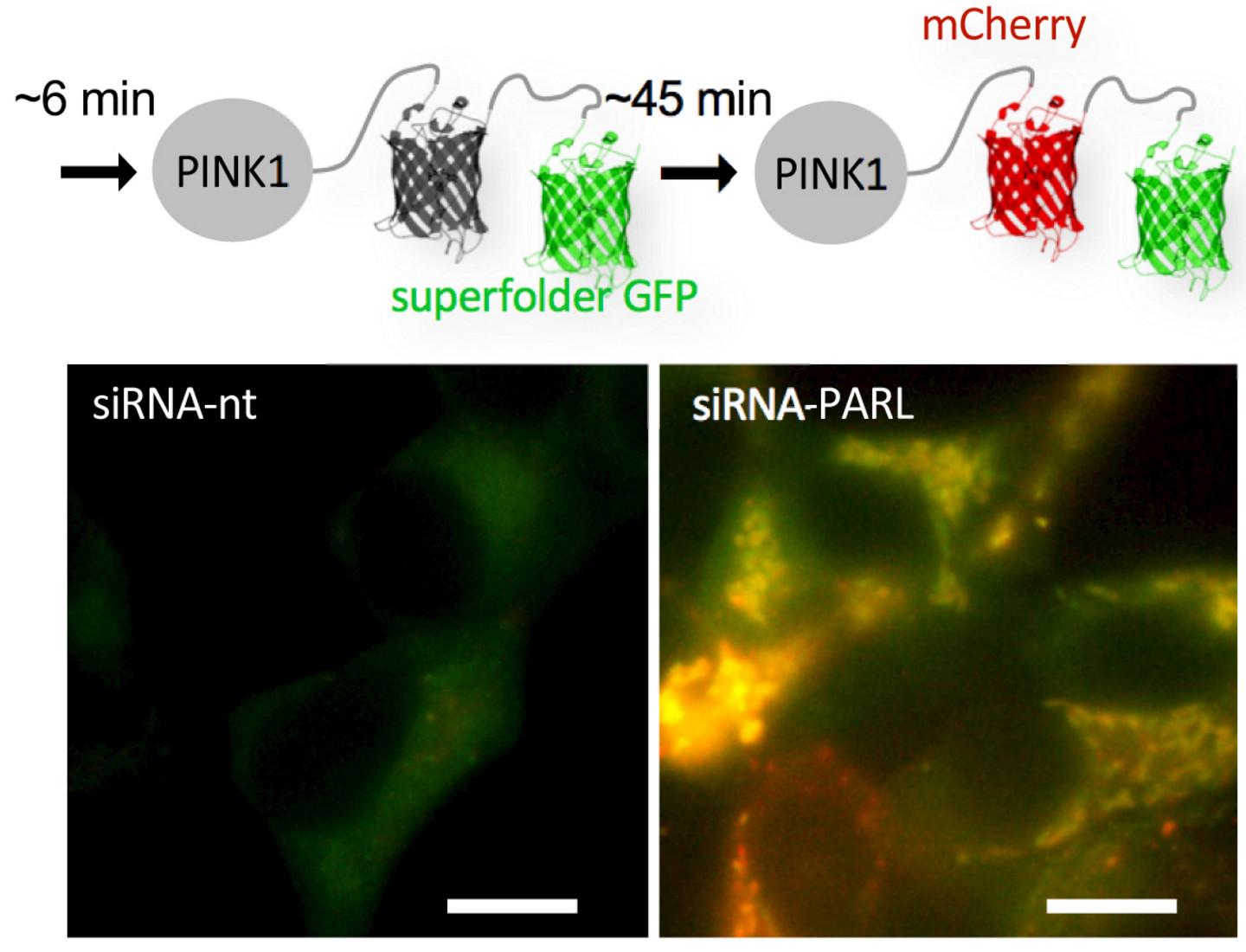
3.) Regulation der mitochondrialen Proteinhomöostase
Mitochondrien sind hochdynamische Organellen, die für zahlreiche essentielle metabolische Prozesse benötigt werden. Mitochondriale Fehlfunktion zeigt gravierende zelluläre Effekte, die mit neurodegenerativen Erkrankungen wie der Parkinson Erkrankung in Zusammenhang stehen. Unsere Arbeit an Mitochondrien begann, als wir entdeckten, dass die Innenmembranprotease PARL den proteasomalen Abbau der Serin/Threonin-Kinase PINK1 auslöst und dadurch diesen zentralen Regulator der Mitophagie steuert. Seit kurzem interessieren wir uns für die Funktion und den Wirkmechanismus der Außenmembran Dislocase Msp1 in Hefe (ATAD1/Thorase beim Menschen). Überraschenderweise konnten wir nachweisen, dass Msp1 mit der ERAD E3 Ubiquitinligase Doa10 zusammenwirkt, um die korrekte Zielfindung C-terminal verankerter Proteine in der äußeren mitochondrialen Membran zu kontrollieren. Unser Hauptinteresse besteht darin, zu verstehen, wie diese Mechanismen der mitochondrialen Proteinhomöostase mit dem proteasomalen und lysosomalen Proteinumsatz interagieren.