Forschungsprojekte
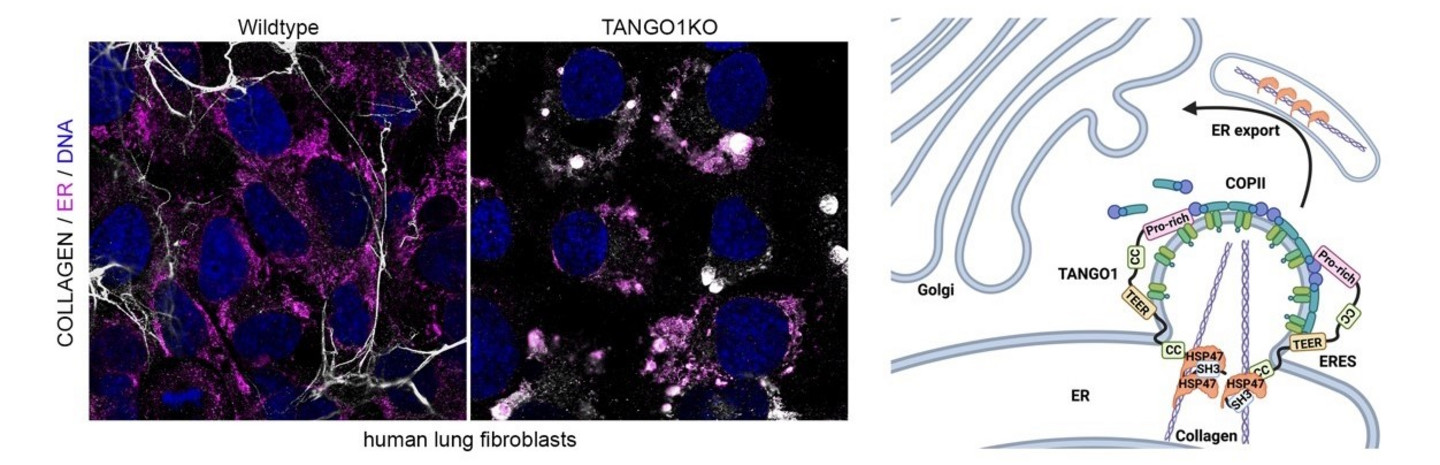
Die Architektur des Gewebes ist dynamisch mit der zellulären Funktion verknüpft, um eine angemessene Gewebehomöostase zu gewährleisten. Die extrazelluläre Matrix (EZM) ist ein entscheidender Bestandteil des Bindegewebes, steht im Zentrum der mechanochemischen Signalübertragung und muss dynamisch in ihrer zell- und gewebetypspezifischen Konfiguration aufgebaut und umgebaut werden. Mutationen in ECM-Proteinen oder Komponenten, die die ECM-Sekretion regulieren, sind bekanntlich die Ursache für schwere Erkrankungen des Bewegungsapparats und altersbedingte Defekte wie schlaffe Haut und brüchige Knochen. Bemerkenswerterweise ist auch die pathologische Überproduktion von ECM, die so genannte Gewebefibrose, häufig eine Folge von altersbedingten Krankheiten und Organversagen, was die Bedeutung einer regulierten Expression und Freisetzung von ECM-Proteinen für die Aufrechterhaltung der Gewebephysiologie und des gesunden Alterns unterstreicht. Wichtig ist, dass die zugrundeliegenden Regulationsmechanismen, die zelluläre Signalwege und die Proteinsekretion miteinander verbinden, nach wie vor weitgehend ungeklärt sind. Trotz jahrzehntelanger Forschung zur (Patho-)Physiologie der extrazellulären Matrix haben sich die meisten Studien bisher auf die extrazelluläre Regulierung des Aufbaus der ECM beschränkt. Die Sekretion von Proteinen der extrazellulären Matrix ist ein streng regulierter Prozess, der zwischen den verschiedenen Kompartimenten des sekretorischen Weges orchestriert werden muss. Darüber hinaus muss die Proteinsekretion dynamisch an Veränderungen in der Gewebestruktur, an veränderte Umweltbedingungen und an mechanische oder zelluläre Belastungen angepasst werden, um die Gewebehomöostase zu gewährleisten. Nach der Synthese im endoplasmatischen Retikulum (ER) werden sekretierte sperrige Frachtproteine wie ECM-Proteine vom einzigartigen Cargorezeptor TANGO1 an ER-Exit-Sites (ERES) erkannt. Dies ist ein essentieller Schritt für den ordnungsgemäßen Export aus dem ER. TANGO1 ist ein in der ER-Membranständiges Protein, das Cargos im ER-Lumen bindet und COPII-Komponenten für den Export von Cargos aus ERES auf der zytosolischen Seite der ER Membran rekrutiert. Die molekularen Mechanismen, die die Auswahl der Cargos steuern, sind jedoch nach wie vor kaum verstanden.
Das übergeordnete Ziel meiner Arbeitsgruppe ist es, herauszufinden, wie die Sekretion, der Aufbau und die Homöostase der EZM in gesunden Zellen reguliert werden und wie die Dysregulation dieser Maschinerie die Gewebsarchitektur in der menschlichen Entwicklung, bei Krankheiten und während des Alterns verändert.